I.2 Fotosyntes, nödvändiga förutsättningar för det
Fotosyntes i gröna växter är processen att omvandla ljus till kemisk energi från organiska föreningar syntetiserade från koldioxid och vatten. Processen för fotosyntes är en kedja av redoxreaktioner, vars helhet är uppdelad i två faser - ljus och mörk.
1. Lätt fas. Denna fas kännetecknas av det faktum att energin från solstrålning som absorberas av pigmenten i kloroplastsystemet omvandlas till elektrokemisk energi.
När ljus verkar på kloroplasten börjar ett elektronflöde genom ett system av bärare - komplexa organiska föreningar inbyggda i tylakoidmembranen. Överföringen av elektroner längs ETC är associerad med det aktiva flödet av protoner genom tylakoidmembranet från stroma in i tylakoiden. I tylakoidutrymmet ökar koncentrationen av protoner på grund av splittring av vattenmolekyler och som ett resultat av oxidation av elektronbäraren plastokinon på insidan av membranet. När protoner går tillbaka längs gradienten från tylakoidutrymmet till stroma syntetiseras ATP på tylakoidens yttre yta med deltagande av enzymet ATP-syntetas från ADP och fosforsyra, det vill säga fotosyntetisk fosforylering sker med lagring av energi i ATP, som sedan passerar in i kloroplastens stroma.
Överföringen av elektroner slutar enligt följande. Efter att ha nått den yttre ytan av tylakoidmembranet, följer ett par elektroner med en vätejon belägen i stroman. Både elektroner och vätejonen är fästa till vätebärarmolekylen – NADP+ (nicatinamidadenin dinukleotid phosphate), som omvandlas till sin reducerade form
NADP H+H+:
NADP++2Н++2е-→NADP H+H+
Följaktligen används elektroner aktiverade av ljusenergi för att fästa väteatomen till dess bärare, d.v.s. för att reducera NADP+ V NADP H+H+, som från den yttre ytan av det fotosyntetiska membranet passerar in i stroma.
I klorofyllmolekyler som har förlorat sina elektroner fungerar de resulterande elektron-"hålen" som ett starkt oxidationsmedel och tar bort elektroner från vattenmolekyler. Genom en serie bärare överförs dessa elektroner till klorofyllmolekylen och fyller "hålet". Fotooxidation (fotolys) av vatten sker inuti tylakoiden, som ett resultat av vilket fritt syre frigörs och vätejoner ackumuleras
2H2O^4H++4e-+02
Under fotosyntesens lätta fas inträffar således tre processer: bildandet av syre på grund av nedbrytningen av vatten, syntesen av ATP och bildandet av väteatomer i form av NADP H2. Syre diffunderar in i atmosfären och ATP och NADP H2 transporteras in i plastidmatrisen och deltar i mörkfasprocessen.
2. Mörk fas fotosyntesen sker i kloroplastmatrisen både i ljus och mörker och representerar en serie sekventiella omvandlingar av CO2 som kommer från luften. Mörkfasreaktioner utförs med hjälp av energin från ATP och NADP H2 och användning av sockerarter med fem kolatomer som finns i plastider, varav en, ribulosdifosfat, är en CO2-acceptor. Enzymer kombinerar sockerarter med fem kolatomer med koldioxid i luften. I detta fall bildas föreningar som successivt reduceras till en glukosmolekyl med sex kol.
Total reaktion av fotosyntes
6СО2+6Н2 ljusenergi С6Н12О6+6О2
Klorofyll
Under fotosyntesprocessen, förutom monosackarider (glukos, etc.), som omvandlas till stärkelse och lagras av växten, syntetiseras monomerer av andra organiska föreningar - aminosyror, glycerol och fettsyror. Således, tack vare fotosyntesen, förser växtceller, eller mer exakt, klorofyllinnehållande celler, sig själva och alla levande varelser på jorden med de nödvändiga organiska ämnena och syre.
I.3 Celldelning
Tre metoder för delning av eukaryota celler har beskrivits: amitos (direkt delning), mitos (indirekt delning) och meios (reduktionsdelning).
Amitos- en relativt sällsynt metod för celldelning. Vid amitos delas interfaskärnan genom förträngning, och enhetlig fördelning av det ärftliga materialet säkerställs inte. Ofta delar sig kärnan utan efterföljande separation av cytoplasman och binukleära celler bildas. En cell som har genomgått amitos kan därefter inte komma in i den normala mitotiska cykeln. Därför förekommer amitos som regel i celler och vävnader som är dömda till döden.
Mitos. Mitos, eller indirekt delning, är den huvudsakliga metoden för delning av eukaryota celler. Mitos är uppdelningen av kärnan, vilket leder till bildandet av två dotterkärnor, som var och en har exakt samma uppsättning kromosomer som fanns i moderkärnan.
I den kontinuerliga processen med mitotisk division finns det fyra faser: profas, metafas, anafas och telofas.
Prophase– den längsta fasen av mitos, när hela kärnans struktur genomgår omstrukturering för delning. I prophase förkortas och förtjockas kromosomerna på grund av deras spiralisering. Vid denna tidpunkt är kromosomerna dubbla (fördubbling sker i S-perioden av interfas) och består av två kromatider anslutna till varandra i regionen för den primära förträngningen genom en speciell struktur - centromeren. Samtidigt med förtjockningen av kromosomerna försvinner nukleolen och kärnmembranfragmenten (bryts upp i separata tankar). Efter kollapsen av kärnmembranet ligger kromosomerna fritt och slumpmässigt i cytoplasman. Bildandet av den akromatiska spindeln börjar - fissionsspindeln, som representerar ett system av trådar som kommer från cellens poler. Spindelfilamenten har en diameter på ca 25 nm. Dessa är buntar av mikrotubuli som består av subenheter av proteinet tubulin. Mikrotubuli börjar bildas från centriolen eller från kromosomerna (i växtceller).
Metafas. I metafas fullbordas bildandet av delningsspindeln, som består av två typer av mikrotubuli: kromosomala, som binder till kromosomernas centromerer, och centrosomala (polära), som sträcker sig från pol till pol i cellen. Varje dubbelkromosom är fäst vid spindelns mikrotubuli. Kromosomer verkar tryckas ut av mikrotubuli in i cellens ekvatorregion, d.v.s. placerade på lika avstånd från polerna. De ligger i samma plan och bildar den så kallade ekvatorial- eller metafasplattan. I metafas är kromosomernas dubbla struktur tydligt, ansluten endast vid centromeren. Det är under denna period som det är lätt att räkna antalet kromosomer och studera deras morfologiska egenskaper.
Anafas börjar med att dividera centromeren. Varje kromatid av en kromosom blir en oberoende kromosom. Sammandragning av dragfilamenten i akromatinspindeln för dem till cellens motsatta poler. Som ett resultat har varje pol i cellen samma antal kromosomer som det fanns i modercellen, och deras uppsättning är densamma.
Telofas – sista fasen av mitos. Kromosomerna despirerar och blir dåligt synliga. Vid varje pol återskapas ett kärnhölje runt kromosomerna. Nukleoler bildas, spindeln försvinner. I de resulterande kärnorna består nu varje kromosom av endast en kromatid, snarare än två.
Mörkreaktioner som uppstår i stroma kräver inte ljus. Minskningen av CO 2 sker på grund av energi (ATP) och reducerande kraft (NADPH 2) som genereras under ljusreaktioner. Mörkreaktioner styrs av enzymer. Sekvensen av dessa reaktioner bestämdes i USA av Calvin, Benson och Bassem mellan 1946 och 1953; 1961 tilldelades Calvin Nobelpriset för detta arbete.
Calvins experiment
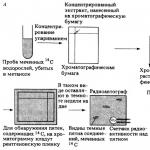
Calvins arbete baserades på användningen av den radioaktiva isotopen kol 14 C (halveringstid 5570 år, se bilaga 1.3), som blev tillgänglig för forskare först 1945. Dessutom använde Calvin papperskromatografi, som vid den tiden var relativt sett ny, ännu inte lite vanlig metod. Kulturer av encelliga grönalger Chlorella (Chlorella) odlades i en speciell apparat (Fig. 9.17). Kulturen hölls vid 14 CO2 under olika tidsperioder, sedan fixerades cellerna snabbt genom att hälla suspensionen i varm metanol. Lösliga fotosyntetiska produkter extraherades, koncentrerades och separerades med användning av tvådimensionell papperskromatografi(Fig. 9.18 och Bilaga 1.8.2). Målet var att spåra vägen genom vilken märkt kol passerar (genom en serie mellanprodukter) in i fotosyntesens slutprodukter. Placeringen av radioaktiva föreningar på papper bestämdes med användning av autoradiografi: för att göra detta placerades fotografisk film känslig för 14 C-strålning på kromatogrammet, och den exponerades, det vill säga svärtades, på de platser där radioaktiva ämnen fanns (fig. 9.18). Inom bara en minut efter inkubation med 14 CO 2 syntetiserades många sockerarter och organiska syror, inklusive olika aminosyror. Men Calvin kunde med mycket korta exponeringar - i 5 sekunder eller mindre - identifiera den första produkten av fotosyntesen och fastställa att det var en syra som innehöll tre kolatomer, nämligen fosfoglycerinsyra(FGK). Han räknade sedan ut hela kedjan av mellanprodukter genom vilken det fixerade kolet överförs; dessa stadier kommer att diskuteras senare. Sedan dess har dessa reaktioner kallats Calvin cykel(eller Calvin-Benson-Bassem-cykeln).

Ris. 9.18. A. Fixering av 14 CO 2 i alger under kortvarig belysning. Bestämning av fixeringsprodukter med papperskromatografi och autoradiografi. B. Autoradiografier av fotosyntesprodukter erhållna efter korttidsbelysning av alger i närvaro av 14 CO 2
9.18. Vilka är fördelarna med att använda långlivade radioaktiva isotoper i biologisk forskning?
9.19. Vilka fördelar kan du få genom att ta chlorella istället för en högre växt?
9.20. Varför är Calvin-apparatkärlet platt och inte sfäriskt?
Carbon Pathway Etapper
Koldioxidfixering:

CO 2 -acceptorn är ett socker med fem kolatomer (pentos) ribulosbisfosfat(dvs ribulos med två fosfatgrupper; denna förening kallades tidigare ribulosdifosfat). Tillsatsen av CO 2 till ett visst ämne kallas karboxylering, och enzymet som katalyserar en sådan reaktion är karboxylas. Den resulterande sexkolsprodukten är instabil och bryts omedelbart ner i två molekyler fosfoglycerinsyra(FGK), som är den första produkten av fotosyntesen. Enzymet ribulosbisfosfatkarboxylas finns i stora mängder i kloroplasternas stroma - det är faktiskt det vanligaste proteinet i världen.
Återhämtningsfas:

FHA innehåller tre kolatomer och har en sur karboxylgrupp (-COOH). TP är triosfosfat eller glyceraldehydfosfat (trekolssocker); den har en aldehydgrupp (-CHO).
För att avlägsna syre från PGA (dvs för att återställa det), används den reducerande kraften hos NADPH 2 och energin hos ATP. Reaktionen fortskrider i två steg: först förbrukas en del av ATP som bildas under ljusreaktioner, och sedan används all NADP·H 2, som också erhålls i ljus. Det totala resultatet är reduktionen av karboxylgruppen i syran (-COOH) till aldehydgruppen (-CHO). Reaktionsprodukten är triosfosfat, det vill säga ett trekolssocker med en fosfatgrupp bunden till den. Denna förening har mer kemisk energi än PGA och är den första kolhydraten som produceras av fotosyntes.
Regenerering av acceptorn för CO 2 - ribulosbisfosfat. En del av triosfosfatet (TP) måste spenderas på regenereringen av ribulosbisfosfat, som används i den första reaktionen. Denna process är en komplex cykel som involverar sockerfosfater med 3, 4, 5, 6, 7 kolatomer. Det är här resten av ATP:n förbrukas. Alla mörka reaktioner sammanfattas i fig. 9.19. I denna figur avbildas Calvincykeln som en "svart låda", i vilken CO 2 och H 2 O kommer in på ena sidan och triosfosfat kommer ut på den andra sidan. Som kan ses från detta diagram används ATP-resten för att fosforylera ribulosbisfosfat, men detaljerna i denna komplexa reaktionskedja visas inte.

Från fig. 9.19 kan vi härleda följande sammanfattande ekvation:

Det är viktigt att notera här att bildningen av två molekyler triosfosfat kräver sex molekyler CO 2. Ekvationen kan förenklas genom att dividera alla koefficienter med 6:

9.21. Rita om figuren. 9.19, som endast anger antalet kolatomer som deltar i reaktionerna; till exempel, istället för 6 RiBF skriv "6 × 5C", etc.
Grundläggande information om processen för fotosyntes sammanfattas i tabell. 9.6.
| Lätta reaktioner | Mörka reaktioner | |
| Lokalisering i kloroplaster | Tylakoider | Stroma |
| Reaktioner | Fotokemiska, dvs de kräver ljus. Ljusenergi orsakar överföring av elektroner från "donatorer" av elektroner till deras "acceptorer" i antingen en icke-cyklisk eller cyklisk väg. Två fotosystem är inblandade - Ι och ΙΙ. De innehåller klorofyllmolekyler som, när de absorberar ljusenergi, avger elektroner. Vatten fungerar som en elektrondonator för den icke-cykliska vägen. Elektronöverföring leder till bildandet av ATP (fotofosforylering) och NADPH 2 (se även tabell 9.5). | De kräver inget ljus. CO 2 fixeras när den binder till en acceptor med fem kolatomer, ribulosbisfosfat (RiBP); i detta fall bildas två molekyler av trekolföreningen fosfoglycerinsyra (PGA), den första produkten av fotosyntesen. Ett antal reaktioner inträffar, gemensamt kallade Calvin-cykeln; i detta fall regenereras acceptorn för CO 2 -RiBP, och FGA reduceras och omvandlas till socker (se även Fig. 9.19). |
| Kombinerade ekvationer |
Adenosintrifosforsyra (ATP) är en universell källa och huvudenergiackumulator i levande celler. ATP finns i alla växt- och djurceller. Mängden ATP är i genomsnitt 0,04% (av cellens våta vikt), den största mängden ATP (0,2-0,5%) finns i skelettmuskulaturen. I en cell förbrukas en ATP-molekyl inom en minut från dess bildande. Hos människor produceras och förstörs en mängd ATP som är lika med kroppsvikten var 24:e timme.
ATP är en mononukleotid som består av kvävebasrester (adenin), ribos och tre fosforsyrarester. Eftersom ATP inte innehåller en, utan tre fosforsyrarester, tillhör den ribonukleosidtrifosfater.
Det mesta av arbetet som sker i celler använder energin från ATP-hydrolys. I det här fallet, när den terminala resten av fosforsyra elimineras, omvandlas ATP till ADP (adenosindifosforsyra), och när den andra fosforsyraresten elimineras, förvandlas den till AMP (adenosinmonofosforsyra). Det fria energiutbytet vid eliminering av både den terminala och andra resten av fosforsyra är cirka 30,6 kJ/mol. Elimineringen av den tredje fosfatgruppen åtföljs av frisättningen av endast 13,8 kJ/mol. Bindningarna mellan den terminala och andra, andra och första fosforsyraresterna kallas makroergisk(hög energi).
ATP-reserverna fylls på hela tiden. I alla organismers celler sker ATP-syntes i processen fosforylering, dvs. tillsats av fosforsyra till ADF. Fosforylering sker med varierande intensitet under andning (mitokondrier), glykolys (cytoplasma) och fotosyntes (kloroplaster).
ATP är huvudlänken mellan processer som åtföljs av frigöring och ackumulering av energi och processer som sker med energiförbrukning. Dessutom är ATP, tillsammans med andra ribonukleosidtrifosfater (GTP, CTP, UTP), ett substrat för RNA-syntes.
Förutom ATP finns det andra molekyler med makroerga bindningar - UTP (uridintrifosforsyra), GTP (guanosintrifosforsyra), CTP (cytidintrifosforsyra), vars energi används för biosyntes av protein (GTP), polysackarider (UTP), fosfolipider (CTP). Men alla av dem bildas på grund av energin från ATP.
Förutom mononukleotider spelar dinukleotider (NAD +, NADP +, FAD), tillhörande gruppen av koenzymer (organiska molekyler som behåller kontakten med enzymet endast under reaktionen), en viktig roll i metaboliska reaktioner. NAD+ (nikotinamidadenindinukleotid), NADP+ (nikotinamidadenindinukleotidfosfat) är dinukleotider som innehåller två kvävehaltiga baser - adenin och nikotinsyraamid - ett derivat av vitamin PP), två ribosrester och två fosforsyrarester. Om ATP är en universell energikälla, då NAD + och NADP + är universella acceptorer, och deras återställda former är NADH Och NADPH – universella givare reduktionsekvivalenter (två elektroner och en proton). Kväveatomen som ingår i nikotinsyraamidresten är fyrvärd och har en positiv laddning ( NAD +). Denna kvävebas fäster lätt två elektroner och en proton (dvs. den reduceras) i de reaktioner där, med deltagande av dehydrogenasenzymer, två väteatomer avlägsnas från substratet (den andra protonen går i lösning):
Substrat-H2 + NAD + substrat + NADH + H+
I omvända reaktioner oxiderar enzymer NADH eller NADPH, reducera substrat genom att lägga till väteatomer till dem (den andra protonen kommer från lösningen).
FAD – flavinadenindinukleotid– ett derivat av vitamin B2 (riboflavin) är också en kofaktor för dehydrogenaser, men FLUGA lägger till två protoner och två elektroner, vilket reducerar till FADN 2.
NADH - grunden för energi och liv
I sin vanliga mening kan biologiskt liv definieras som förmågan att generera energi i en cell. Denna energi är högenergifosfatbindningar av kemikalier som syntetiseras i kroppen. De viktigaste högenergiföreningarna är adenosintrifosfat (ATP), guanosintrifosfat (GTP), kreatinfosforsyra, nikotinamid-dinukleotidfosfat (NAD(H) och NADP(H)), fosforylerade kolhydrater.
Nikotinamidadenindinukleotid (NADH) är ett koenzym som finns i alla levande celler och är en del av dehydrogenasgruppen av enzymer som katalyserar redoxreaktioner; utför funktionen av en bärare av elektroner och väte, som den tar emot från oxiderbara ämnen. Den reducerade formen (NADH) kan överföra dem till andra ämnen.
Hur man förbättrar prestandan
Vad är NADH? Många människor kallar det "en förkortning för livet." Och det är det verkligen. NADH (nikotinamidadenindinukleotidkoenzym) finns i alla levande celler och är ett viktigt element genom vilket energi produceras inuti celler. NADH är involverad i produktionen av ATP (ATP). NAD(H), som en universell energimolekyl, till skillnad från ATP, kan ständigt avlasta mitokondrier från överdriven ackumulering av laktat mot bildandet av pyruvat från det, på grund av stimulering av pyruvatdehydrogenaskomplexet, som är specifikt känsligt för NAD(H) /NAD-förhållande.
Kroniskt trötthetssyndrom: Fokus på mitokondrier
Ett antal kliniska studier har visat effektiviteten av NADH-läkemedel vid CFS. Den dagliga dosen var vanligtvis 50 mg. Den mest kraftfulla effekten inträffade efter 2-4 veckors behandling. Tröttheten minskade med 37-52%. Dessutom förbättrades en sådan objektiv kognitiv parameter som koncentration av uppmärksamhet.
NADH vid behandling av kroniskt trötthetssyndrom
NADH (vitamin B3-koenzym), som finns i alla levande celler, är en del av dehydrogenasgruppen av enzymer som katalyserar redoxreaktioner; utför funktionen av en bärare av elektroner och väte, som den tar emot från oxiderbara ämnen. Det är en reservkälla för energi i celler. Det deltar i nästan alla energiproduktionsreaktioner, vilket säkerställer cellandning. Genom att påverka motsvarande processer i hjärnan kan vitamin B3-koenzym förhindra nervcellers död vid hypoxi eller åldersrelaterade förändringar. Deltar i avgiftningsprocesser i levern. Nyligen har dess förmåga att blockera laktatdehydrogenas och därigenom begränsa ischemisk och/eller hypoxisk skada på myokardiet fastställts. Studier av effektiviteten av oral administrering vid behandling av kroniskt trötthetssyndrom har bekräftat dess aktiverande effekt på människors tillstånd.
NADH i sport och medicin: granskning av utländsk litteratur
Vi skrev om NADH (nikotinamidadenindinukleotidfosfat) i tidigare artiklar. Nu vill vi ge information från engelskspråkiga källor om denna substanss roll och betydelse för energiomsättningen i kroppen, dess effekt på nervsystemet och dess roll i utvecklingen av ett antal patologiska situationer och möjligheter för användning i medicin och sport. (Ladda ner monografi om NADH).
Herbalife Quickspark CoEnzyme 1 (NADH) ATP-energi
Naturlig energi på cellnivå
Quickspark är en produkt från företaget Herbalife. Det är en stabil form av Vitamin B3 CoEnzyme1. CoEnzyme1 hittades 1906 i Österrike av en vetenskapsman som heter professor George Birkmayer. CoEnzyme1 utvecklades för medicinska ändamål och användes under andra världskriget.
NADH (Enada)
Nikotinamidadenindinukleotid (NADH) är ett ämne som hjälper enzymernas funktion i kroppen. NADH spelar en roll i produktionen av energi och hjälper till att producera L-dopa, som kroppen omvandlar till signalsubstansen dopamin. NADH utvärderas för många tillstånd och kan vara till hjälp för att förbättra mental funktionalitet och minne.
Fotosyntes är syntesen av organiska ämnen från koldioxid och vatten med obligatorisk användning av ljusenergi: 6CO 2 +6H 2 O + Q ljus →C 6 H 12 O 6 +6O 2. Fotosyntes är en komplex process i flera steg; Fotosyntesreaktioner är indelade i två grupper: ljusfasreaktioner och mörkfasreaktioner.
Lätt fas. Förekommer endast i närvaro av ljus i tylakoidmembran med deltagande av klorofyll, elektrontransportproteiner och enzymet ATP-syntetas. Under påverkan av ett ljuskvantum exciteras klorofyllelektroner, lämnar molekylen och går in på utsidan av tylakoidmembranet, som i slutändan blir negativt laddat. Oxiderade klorofyllmolekyler reduceras och tar elektroner från vatten som finns i det intratylakoida utrymmet. Detta leder till nedbrytning och fotolys av vatten: H 2 O+ Q ljus →H + +OH - . Hydroxidjoner ger upp sina elektroner och förvandlas till reaktiva radikaler ∙OH: OH - →∙OH+e - . ∙OH-radikaler kombineras för att bilda vatten och fritt syre: 4HO∙→ 2H 2 O+O 2. I det här fallet avlägsnas syre till den yttre miljön och protoner ackumuleras inuti tylakoiden i "protonreservoaren". Som ett resultat är tylakoidmembranet å ena sidan positivt laddat på grund av H +, och å andra sidan, på grund av elektroner, laddas det negativt. När potentialskillnaden mellan de yttre och inre sidorna av tylakoidmembranet når 200 mV, trycks protoner genom ATP-syntetaskanalerna och ADP fosforyleras till ATP; atomärt väte används för att återställa den specifika bäraren NADP + till NADP∙H 2: 2H + +2 e - + NADP→ NADP∙H 2. I ljusfasen sker alltså fotolys av vatten, vilket åtföljs av tre viktiga processer: 1) ATP-syntes; 2) bildning av NADP∙H2; 3) bildning av syre. Syre diffunderar in i atmosfären, ATP och NADP∙H 2 transporteras in i kloroplastens stroma och deltar i den mörka fasens processer.
Mörk fas. Förekommer i kloroplastens stroma. Dess reaktioner kräver ljusenergi, så de förekommer inte bara i ljuset utan också i mörkret. De mörka fasreaktionerna representerar en kedja av successiva omvandlingar av koldioxid (från luften), vilket leder till bildandet av glukos och andra organiska ämnen. Först sker CO 2 -fixering, acceptorn är sockerribulosbifosfatet, katalyserat av ribulosbifosfatkarboxylas. Som ett resultat av karboxylering av ribulosbifosfat bildas en instabil sexkolsförening, som omedelbart bryts ner i två molekyler fosfoglycerinsyra. Sedan uppstår en cykel av reaktioner där PGA genom en serie mellanprodukter omvandlas till glukos. Energin från ATP och NADPH 2 som bildas i ljusfasen används. (Calvin-cykeln).
23. Co2-assimileringsreaktioner i fotosyntesens mörka fas.
Calvincykeln är huvudvägen för CO 2 -assimilering. Dekarboxyleringsfas - koldioxid binder med ribulosbifosfat för att bilda två fosfoglyceratmolekyler. Denna reaktion katalyseras av ribulosbifosfatkarbosylas.